Класс Ленточные черви: общая характеристика, представители. Половая система ленточных червей Строение размножение и развитие ленточных червей
Выделительная система - протонефридиального типа ( рис. 157 , рис. 158). По бокам вдоль всего тела, непосредственно кнутри от нервных стволов идут два главных выделительных канала. Они начинаются на заднем конце тела, затем направляются кпереди, достигают головки, заворачиваются назад и вновь доходят до заднего конца, заканчиваясь общим выделительным отверстием. Вследствие этого часто получается впечатление, что у цестод имеются 4 продольных канала, тогда как их всего 2, но петлеобразно изогнутых у переднего конца тела. При соединении на заднем конце тела оба канала нередко образуют небольшой общий сократимый мочевой пузырь. У членистых цестод боковые каналы как в головке, так и у заднего края члеников соединяются при помощи поперечных перемычек; выделительная система получает вид лестницы. Когда последний членик цепи отвалится (вследствие периодического отрывания задних кусков стробилы), нового мочевого пузыря уже не образуется, и каждый боковой канал открывается теперь наружу особым отверстием. Многочисленные веточки каналов пронизывают паренхиму и на концах своих замыкаются звездчатыми клетками с мерцательным пламенем.
1. Нервная система.
Имеет некоторые черты централизации. В головной части червя содержится парный нервный узел (ганглий). Нервные элементы собираются в два нервных ствола, идущих вдоль всего тела. Между собой стволы соединяются комиссурами (пучками нервных волокон). От стволов отходят тонкие ветви, образующие нервное сплетение.
2. Кровеносная система.
Ленточные черви лишены сердца и сосудов. Это связано с примитивным эволюционным развитием, где функции крови выполняет межклеточная жидкость, которая переносит вещества из разных частей тела. Каждый сегмент получает необходимые для жизнедеятельности продукты за счёт возможности всасывания веществ поверхностными покровами.
3. Дыхательная система.
Также отсутствует.
4. Пищеварительная система.
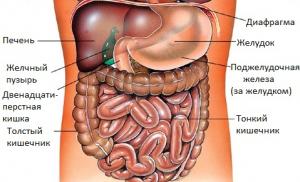
Как таковая отсутствует. Это можно объяснить средой обитания ленточных червей: в кишечнике хозяина (например, человека) у цестод нет потребности перерабатывать пищу, она и так находится в готовом для усвоения виде. Поэтому черви не имеют пищеварительных органов и ферментов, а пищу получают путем диффузии через кожные покровы.
5. Выделительная система.
Протонефридиального типа, что подразумевает наличие большого количества клеток с мерцательным эпителием, связанных с трубочками, образующими сеть. Канальцы сети впадают в крупные каналы, идущие вдоль тела (обычно их два). Два выделительных канала начинаются в терминальном конце туловища цестод, проходят по всему телу, и, собрав жидкость, возвращаются в концевой отдел. Заканчиваются общей выделительной порой.
6. Половая система.
Из всех систем органов самая развитая. Все особи ленточных червей являются гермафродитами. Каждый членик содержит в себе органы обоих полов. Мужская система образована множеством семенников и семявыносящих канальцев, открывающихся в семяпровод. Последний заканчивается семяизвергательным каналом. Женские органы размножения: матка, яичник, яйцеводы, желточники, влагалище. Мужские и женские половые органы открываются в половую клоаку. Оплодотворение происходит между члениками одной особи или между двумя червями.
Общая характеристика класса Cestoda
Строение цестод
Тело обыкновенно бывает сильно вытянутым в длину, лентовидным и в большинстве случаев поделено на значительное число члеников, или проглоттид. Редко тело бывает цельным, нерасчлененным. Передний конец образует небольшую головку, или сколекс, за которым следует нерасчлененная шейка, а за ней идут проглоттиды.
Головка несет органы прикрепления, построенные по типу присосок или по типу крючков. Присоски имеются всегда, тогда, как крючья являются менее постоянным их дополнением. Присоски устроены, в общем, так же, как у сосальщиков, и чаще всего имеются в числе четырех по краю переднего конца головки. Реже вместо типичных присосок головка снабжена двумя вытянутыми в продольном направлении щелевидными присасывательными ямками. Крючья помещаются или прямо на поверхности головки, или на особом переднем выпячивании ее - хоботке, образуя один или несколько венчиков. Хоботок втяжной. В редких случаях (Tetrarhynchus) имеется 4 хоботка, длинных, усаженных многочисленными крючками и способных глубоко вворачиваться в особые влагалища.
Проглоттиды имеют обычно четырехугольную форму, причем число их варьирует от 3 шт. до нескольких тысяч. Передние членики - самые маленькие, по направлению же назад размеры их постепенно возрастают. Во все время жизни червя происходит рост и увеличение числа члеников. Рост идет в области шейки: она удлиняется, и от заднего конца ее отшнуровываются все новые членики. Таким образом, самые молодые членики занимают переднюю часть тела; чем дальше от сколекса расположен членик, тем он старее. Все тело называют цепочкой или стробилой (последнее за сходство с процессом стробиляции сцифостом сцифоидных медуз).
Цестоды обладают типичным кожно-мускульным мешком. Их покровы очень сходны с таковыми у трематод и турбеллярий и построены по тому же типу, что и погруженный эпителий турбеллярий. Тегумент цестод слагается из безъядерного цитоплазматического слоя, при помощи тонких тяжей соединенного с погруженными участками цитоплазмы, несущими ядра.
Отличительная особенность покровов цестод состоит в том, что на поверхности наружного цитоплазматического слоя имеется бесчисленное множество волосковидных выростов (микротрихий), по-видимому, играющих роль в процессе питания. Рассмотреть их можно только с помощью электронного микроскопа. Их ультраструктура принципиально отличается от таковой микроворсинок, характерных для спороцист трематод.
Непосредственно под базальной мембраной располагается 2 слоя: наружный слой кольцевых и внутренний - продольных мускульных волокон. Часто к этим двум слоям присоединяется еще третий, более глубокий слой – слой кольцевых мышечных волокон. Кроме того, имеется система спинно-брюшных мышечных пучков, пронизывающих паренхиму. В последней у большинства цестод разбросаны микроскопические округлые конкреции углекислой извести - так называемые известковые тельца.
Их происхождение и функция до конца не выяснены, но имеется предположение, что известковые тельца возникают как экскреты и выполняют задание своеобразной буферной системы, предохраняющей ленточных червей от вредного влияния кислой среды (например, при миграции личиночных стадий через желудок хозяина).
В паренхиме ленточных червей откладывается значительное количество гликогена, в результате анаэробного расщепления которого цестоды (подобно трематодам) получают энергию, необходимую для жизнедеятельности.
Нервная система цестод
Центральная нервная система состоит из парного мозгового узла, лежащего в головке и посылающего от себя назад несколько пар нервных стволов, соединенных поперечными перемычками (ортогон). Два ствола, расположенных по бокам тела, развиты сильнее остальных. От стволов отходят тонкие веточки, образующие под кожей довольно густое нервное сплетение.
Пищеварительная система цестод
Выделительная система цестод
Выделительная система - протонефридиального типа. По бокам вдоль всего тела, непосредственно внутрь от нервных стволов идут два главных выделительных канала. Они начинаются на заднем конце тела, затем направляются кпереди, достигают головки, заворачиваются назад и вновь доходят до заднего конца, заканчиваясь общим выделительным отверстием. Вследствие этого часто получается впечатление, что у цестод имеются 4 продольных канала, тогда как их всего 2, но петлеобразно изогнутых у переднего конца тела. При соединении на заднем конце тела оба канала нередко образуют небольшой общий сократимый мочевой пузырь. У членистых цестод боковые каналы, как в головке, так и у заднего края члеников соединяются при помощи поперечных перемычек; выделительная система получает вид лестницы. Когда последний членик цепи отвалится (вследствие периодического отрывания задних кусков стробилы), нового мочевого пузыря уже не образуется, и каждый боковой канал открывается теперь наружу особым отверстием. Многочисленные веточки каналов пронизывают паренхиму и на концах своих замыкаются звездчатыми клетками с мерцательным пламенем.
Половая система ленточных червей
Половая система ленточных червей гермафродитная и, в общем, напоминает таковую сосальщиков. Лишь у некоторых из нерасчлененных цестод (Caryophyllaeus) половой аппарат одиночен. У других, например у ремнецов, имеется продольный ряд половых аппаратов, тогда как у членистых цестод в каждой проглоттиде развивается своя половая система.
Отдельные части полового аппарата у разных ленточных червей довольно сильно варьируют, так что для конкретности рассмотрим частный случай бычьего, или невооруженного, солитера (Taeniarhynchus saginatus).
В молодых передних члениках стробилы половые органы еще не развиты и начинаются приблизительно лишь с 200 членика. В следующих члениках с вполне развитой половой системой мужской отдел последней состоит из многочисленных семенников, разбросанных в паренхиме. Тонкие семявыносящие протоки семенников соединяются вместе и образуют общий семяпровод. Последний направляется к одной из узких боковых граней тела и там пронизывает собой совокупительный орган, имеющий вид мускулистой трубки, которая своим концом вдается в глубокую ямку на боковой грани тела - половую клоаку.
Женский отдел системы состоит из ветвистого яичника, проток которого, яйцевод, впадает в оотип, как у трематод. В оотип же поступает и содержимое непарного желточника - сетевидной железы, прилегающей к задней стенке членика. Кроме того, от оотипа отходят два канала. Один, влагалище, тянется рядом с семяпроводом и открывается подле него в половую клоаку. Другой, более широкий, направляется от оотипа вперед по срединной линии членика и заканчивается слепо, это матка.
Яйцеклетки поступают в оотип, куда проникают и спермин через влагалище. В оотипе яйца оплодотворяются, окружаются скорлупой и переводятся в матку, где проходят первую часть своего развития. У цепней вследствие отсутствия выводного отверстия яйца остаются в матке долго и выходят наружу лишь при разрыве стенок членика. Яйца настолько переполняют матку, что последняя сильно разрастается, дает от своего главного ствола много боковых ветвей в обе стороны и занимает значительную часть членика. К этому времени все остальные части половой системы заканчивают свою функцию и подвергаются большей или меньшей атрофии. Членики, в которых осталась лишь сильно разветвленная и набитая яйцами матка, называют "зрелыми". Зрелые членики занимают задний конец цепочки и периодически отрываются целыми группами.
У других цестод может быть ряд разнообразных отступлений. Так, матка, слепо замкнутая у цепня, нередко (например, у Diphyllobothrium и др.) открывается наружу на одной из плоских сторон членика (рис. 160). У таких видов яйца по мере заполнения ими матки выходят из нее в кишечник животного-хозяина. Мужское отверстие и отверстие влагалища могут при этом смещаться на одну из плоских сторон членика. Любопытным изменением является наблюдаемое у некоторых форм частичное или полное удвоение полового аппарата в каждом членике (например, у тыквенного цепня - Dipylidium caninum).
Оплодотворение у ленточных червей происходит как перекрестно, так и посредством самооплодотворения, причем совокупительный орган одного членика вводится во влагалище другого или даже, изгибаясь, во влагалище того же самого членика.
Жизненный цикл цестод
Яйца выходят наружу с испражнениями или посредством разрыва стенок члеников, или вместе с члениками, группы которых периодически отрываются от стробилы. Для дальнейшего развития яйца, как у сосальщиков, должны попасть во вполне определенного промежуточного хозяина, которым для Т. solium служит свинья.
Заражение свиней происходит вследствие их нечистоплотности, так как они охотно роются в отбросах, нечистотах и т. п. У яиц, попавших в кишечник свиньи, скорлупа разрушается, и из яйца выходит личинка - онкосфера, или шестикрючный зародыш - маленький многоклеточный шарик, снабженный шестью хитиноидными крючками. Крючочки эти не соответствуют тем крючкам, которые у многих цестод образуются на сколексе, и отбрасываются при дальнейшем развитии. При помощи крючков онкосфера вбуравливается в стенки желудка или кишки, попадает в лимфатические или кровеносные сосуды и током крови заносится в различные внутренние органы: чаще всего - в печень, мышцы, реже - в легкие, мозг и др. Здесь онкосфера застревает, останавливается и испытывает превращение в стадию финны. Превращение начинается с быстрого роста онкосферы, которая достигает размера крупной горошины. Тело личинки становится полым, представляя собой пузырь, наполненный жидкостью. Это и есть финна, или пузырчатая глиста. Стенки пузыря образуют в одной точке впячивание, на дне которого на внутренней поверхности появляются в виде 4 ямок зачатки присосок, а между ними, на самом дне, венчик небольших крючков. Это впячивание - зачаток головки ленточной глисты, но только ввороченной внутрь пузыря финны.
В теле свиньи дальнейшее развитие финны не имеет места, но финна может несколько лет просуществовать в своем промежуточном хозяине, не погибая. Для достижения половозрелого состояния финна должна попасть в кишечник окончательного хозяина, т. е. человека.
Попав вместе с недостаточно прожаренным или просоленным и т. п. мясом в кишечник человека, финны Т. solium освобождаются из мяса под влиянием пищеварительных соков, а затем головка финны (главным образом под действием желчи) выворачивается наружу, причем крючья и присоски принимают свое нормальное положение. Сморщенный пузырь финны некоторое время висит на конце шейки сколекса в виде хвостового пузыря, а затем отпадает и разрушается. Головка с шейкой начинают усиленно расти и посредством поперечных насечек отшнуровывать на заднем конце последней все новые членики, образуя длинную цепь проглоттид.
Итак, цикл развития Т. solium связан со сменой хозяев и со сложным превращением - онкосферы в финну, а финны в ленточную половозрелую стадию.
У прочих цестод общий ход развития таков же, но отдельные детали, и притом довольно важные, могут варьировать.
Тип строения финны, свойственный свиному солитеру, называется цистицерком. Это небольшой полый пузырь с ввороченной в него одной головкой. Особый род финн, ценур, имеется в тех случаях, когда пузырь финны сильно разрастается и на стенках его вместо одного впячивания образуется множество их, т. е. в одном пузыре закладывается много головок, каждая из которых дает начало половозрелой ленточной особи.
Наконец, особенно сильного развития достигают финны - эхинококки. Пузырь эхинококка вырастает до огромных размеров и образует внутри себя и на стенках много вторичных пузырей меньшей величины - выводковых капсул. На внутренних стенках капсул формируются многочисленные впячивания головок; таким образом, каждая выводковая капсула эхинококка отвечает как бы одному ценуру, а единственный шестикрючный зародыш дает начало нескольким тысячам головок.
Несмотря на внешние различия, все формы финн являются разновидностями одного общего основного типа. По характеру жизненного цикла, представляющего развитие, сопровождающееся метаморфозом, цестоды стоят ближе к моногенеям, чем к трематодам. Последним свойственно чередование поколений типа гетерогонии. Однако у тех видов цестод, развитие которых связано с формированием ценура или эхинококка, жизненный цикл вторично усложняется. В него привходит бесполое размножение пузырчатой стадии, дающей путем почкования множество дочерних головок (сколексов). Имеет место чередование двух поколений - полового и бесполого, т. е. метагенез.
Значение умножения головок состоит (так же как и при размножении редий и спороцист сосальщиков) в увеличении численности потомства и соответственно в усилении возможности заражения окончательного хозяина.
Своеобразный членистый характер тела большинства взрослых лентецов породил несколько отличающихся друг от друга толкований их организации и природы их стробилы.
Теория колониальности цестод
По теории колониальности, или полизоичности, цепочка ленточного червя представляет собой колонию индивидов, образовавшуюся путем бесполого размножения, а именно концевого почкования от одной первичной особи, т. е. сколекса. Каждый членик представляет собой особь такой колонии. При этом цепочка сравнивается со стробилой сцифомедуз, а сколекс - с первичным полипом стробилы - сцифостомой.
В пользу теории колониальности цестод свидетельствуют следующие факты: 1) возможность сравнения с примерами из других групп животных; 2) наличие в каждом членике вполне самостоятельного полового аппарата; 3) значительная независимость от головки, достигаемая члениками некоторых ленточных червей; так, группы набитых яйцами члеников Taeniarhynchus saginatus, оторвавшиеся от цепочки и выведенные наружу, могут ползать еще много часов. Еще большей самостоятельности достигают членики некоторых цестод из кишечника акул (например, Echinobothrium, Trilocularia; рис. 166). Они поодиночке отрываются от стробилы еще незрелыми, долгое время живут в кишечнике хозяина, сильно вырастают за этот период и развивают в себе половые продукты. У таких члеников формируется даже подобие прикрепительного аппарата, при помощи которого они удерживаются в организме хозяина.
Многие исследователи придерживаются теории монозоичности цестод, согласно которой стробила представляет собой одну особь, а членики - лишь участки этой особи.
Действительно, в каждой проглоттиде самостоятелен только половой аппарат, тогда как все другие системы органов говорят об индивидуальности всей цепи в целом, а не отдельных члеников. Так, в нервной системе всего один мозговой ганглий, в выделительной - лишь один мочевой пузырь, и т.д. Наконец, имеется несколько родов цестод, у которых сколекс неясно обособлен от остального тела, содержащего всего один половой аппарат. Такие ленточные черви напоминают трематод. Сторонники теории монозоичности считают подобные формы наиболее примитивными. Причины, вызвавшие умножение числа половых аппаратов и расчленение тела цестод, приводятся различные. В первую очередь, умножение числа половых аппаратов способствует произведению большого числа яиц. Тенденция к этому, быть может, сказывается и в удвоении половых аппаратов в каждом членике, о чем говорилось раньше.
Вероятно, что еще до расчленения тела цестод на проглоттиды в нем образовался продольный ряд половых аппаратов, как у некоторых современных ремнецов (Ligula). Уже позднее соответственно каждому половому аппарату обособился и участок тела в виде членика, все же отвечающий, как видно из его происхождения, не целой особи, а ее части.
К болезням, вызываемым этой группой гельминтов, относятся эхинококкоз, спарганоз и ценуроз.
Для Taenia solium человек может быть как промежуточным, так и окончательным хозяином.
Цестоды имеют лентовидную форму и прикрепляются к слизистой кишечника с помощью присосок или присасывательных щелей, расположенных на головке - сколексе. Позади сколекса находится короткая тонкая шейка, от которой отшнуровываются членики - проглоттиды. По мере созревания очередного членика его оттесняют назад новые, менее зрелые членики. Тело гельминта - стробила - состоит из постоянно удлиняющейся цепочки члеников; их может быть более тысячи, а длина цестод достигает нескольких метров. Созрев, членики начинают выделять яйца.
Когда финнозное мясо попадает в ЖКТ окончательного хозяина, из каждого сколекса развивается взрослый гельминт.
В организме человека обитают такие виды цестод:
Обыкновенный ремнец (Ligula intestinatis), Лентец широкий (Diphyllobothrium latum), Невооруженный цепень (Taeniarhynchus saginatus), Вооруженный цепень (Taenia solium), Цепень карликовый (Hymenolepis nana), Эхинококк (Echinococcus granulosus).
- солитер достигает в длину 2-3 метров, имея лентовидное, белое тело. Промежуточным хозяином такого гельминта являются домашние животные (часто свиньи, чуть менее редко - собаки, кошки, кролики и т.п.), а окончательным - человек. При заражении у человека провоцирует развитие тениоза. Симптоматика данного заболевания разнообразна по своему характеру. Проявляется тошнотой, рвотой, поносом, отсутствием аппетита. Тяжелое осложнение тениоза - цистицеркоз, при котором в организме пациента развиваются финнозные стадии. Для профилактики заболевания человеку не стоит питаться непроверенной, сырой или полусырой свининой, свиным салом, а также следить за личной гигиеной;
- бычий цепень , может достигать в длину до 4-10 метров. Промежуточным хозяином данного гельминта являются дикие и домашние животные (антилопы, крупный и мелкий рогатый скот), а окончательным носителем выступает человек. При заражении гельминтом развивается тениаринхоз. И хотя это заболевание менее опасно, нежели тениоз, однако оно переносится пациентом крайне тяжело и требует длительного лечения. Профилактическими мерами являются покупка сертифицированного мяса, его качественная термообработка, а также соблюдение правил личной гигиены;
- эхинококк вырастает в длину до 3-5 мм. Поражает представителей крупного рогатого скота, овец, свиней и даже человека. В качестве окончательного хозяина для такого гельминта выступают собака, волк, лисица. Особенно яркой симптоматики эхинококкоз не дает, поэтому он может оставаться без внимания пациента и адекватного лечения длительное время. При поражении головного мозга возможны головокружения, головные боли, мигрени, эпилептические припадки;

Отличительные особенности цестод
Ленточные черви являются наиболее крупными по размеру червями, способны стать причиной опасных заболеваний человека.
Строение тела
Лентовидное тело цестоды имеет разнообразные размеры в зависимости от вида микроорганизма и среды его обитания. Максимальная длина ленточного гельминта достигала 12 м. А состоит он из устрашающего вида головы (на ней расположено несколько присосок и крючьев), шейки, а также хвоста (он разделен на множество члеников с яйцами). В зоне шейки гельминт растет, стремительным образом увеличиваясь в размерах.
В процессе роста молодой особи в зоне шеи появляется поперечная перетяжка, которая отделяет заднюю часть гельминта, превращающуюся в проглоттиду. Чем старше становится червь, тем больше члеников он имеет. Новые членики более мелкого размера расположены у головы, а старые (более крупные) поочередно отодвигаются к хвосту, зреют и отрываются от стробилы. Зрелые яйца попадают в окружающую среду и поедаются животными вместе с травой, на которой они закрепляются, или водой, в которую попадают.
Особенности половой системы

Однако половая система у гельминтов ленточного типа не образуется с первых моментов жизни особи. Помимо формы тела, с ростом гельминта меняется и состояние половой системы члеников. Самые молодые представители цестод не имеют половой системы. В средних по зрелости члениках начинают формироваться органы мужской половой системы. В самых зрелых проглоттидах, которые располагаются приблизительно в середине стробилы, начинают свое развитие органы женской половой системы. Таким образом, членик приобретает признаки гермафродита.
Строение мышечной системы
Мышечная система представлена кольцевыми и продольными слоями, пучками дорсовентральных мышц. Кожно-мускульный мешок содержит паренхиму и внутренние органы.
Выделительная и нервная системы
Центральная нервная система представлена главными стволами по бокам выделительных каналов.
Способ питания и дыхания